Embryo v štádiu gastruly, koľko vrstiev má. Embryonálny vývoj. Pozrite sa, čo je „Gastrula“ v iných slovníkoch
Na konci obdobia štiepenia mnohobunkové zvieratá začínajú obdobie tvorby zárodočných vrstiev - gastrulácia. gastrulácia je proces tvorby dvojvrstvového embrya. Podstata spočíva vo vytvorení jednovrstvového embrya - dvojvrstvového, pozostávajúceho z ektodermu a endodermu Embryo vytvorené v dôsledku gastrulácie sa nazýva gastrula.
Blastopore- otvor, ktorým komunikuje dutina primárneho čreva zvieracieho embrya v štádiu gastruly životné prostredie.
Tvorba skorej gastruly sa vyskytuje nasledujúcimi spôsobmi:
Imigrácia (vysťahovanie buniek), v coelenterátoch;
Invaginácia (invaginácia), v lancelete;
Epiboly (znečistenie), v žabe;
Delaminácia (štiepenie), v koelenterátoch.
Keď imigráciačasť blastodermových buniek z povrchu embrya prechádza do blastocoelu. Vonkajšia vrstva je ektoderm a vnútorná vrstva je endoderm. Blastocoel je naplnený bunkami.
S intususcepciou určitá časť blastodermy (vegetatívny pól) sa ohýba dovnútra a dosiahne zvierací pól. Vytvára sa dvojvrstvová embryo-gastrula. Vonkajšia vrstva buniek je ektoderm, vnútorná vrstva je endoderm, ktorý vystiela dutinu primárneho čreva (gastrocoel). Otvor, ktorým dutina komunikuje s vonkajším prostredím, sa nazýva primárne ústa – blastopór. U protostómov (červy, mäkkýše, článkonožce) sa mení na ústny otvor, u deuterostómov na konečník a na opačnom konci sa tvoria ústa (strunatce).
epiboly charakteristické pre zvieratá, ktoré sa vyvíjajú z telolecitálnych vajíčok. Vznik gastruly je spôsobený rýchlym množením mikromérov, ktoré prerastajú vegetačný pól. Makroméry sú vo vnútri embrya. Nedochádza k tvorbe blastoporov a nedochádza k gastrocoelu.
Delaminácia vyskytuje sa v coelenterátoch, ktorých blastula je podobná morule. Bunky blastodermu sú rozdelené na vonkajšie a vnútorné vrstvy. Vonkajšia vrstva tvorí ektoderm, vnútorná vrstva tvorí endoderm
Všetky mnohobunkové organizmy, okrem húb a koelenterátov, majú 3. zárodočnú vrstvu - mezoderm - neurulácia. Tvorba mezodermu prebieha dvoma spôsobmi: teloblastickým alebo enterocelóznym.
Teloblastický metóda je charakteristická pre primárne ústa.Na hranici medzi ektodermou a endodermou na stranách blastoporcely sa teloblasty začínajú deliť a dávajú vznik mezodermu.
Enterocelický metóda je typická pre deuterostómy. Bunky, ktoré tvoria mezoderm, sú izolované vo forme vreciek primárneho čreva. Dutiny vreciek sa menia na celok.Mezoderm je rozdelený na samostatné úseky - somity, z ktorých sa vytvárajú určité tkanivá a orgány.
Organogenéza. Neirula. Záložka somites.
Po vytvorení mezodermu začína proces histo- a organogenézy. Najprv sa vytvoria axiálne orgány - neurálnej trubice, akord, potom všetky ostatné orgány. Vytvára sa neurula.
Organogenéza- kladenie a tvorba orgánov. Primárna organogenéza- uloženie osových orgánov (tetiva, nervová trubica, črevná trubica). sekundárna organogenéza- tvorba všetkých ostatných orgánov.
Tvorba axiálnych orgánov v embryách strunatcov:
1. Ektoderm na dorzálnej strane embrya sa ohýba a vytvára pozdĺžnu ryhu, ktorej okraje sa uzatvárajú. Vytvorená nervová trubica ponorí sa do ektodermy.
2. Postupne sa oddeľuje a formuje dorzálna časť endodermu, ktorá sa nachádza pod nervovým púčikom akord.
3. Z ektodermy a endodermy vzniká črevná trubica.
ektodermu- epidermis, kožné žľazy, vlasy, sklovina, spojivka, šošovka, sietnica, uši, epiteliálna výstelka nosnej dutiny a dutiny ústnej, konečník a vagína, predná a zadná hypofýza, CNS, dreň nadobličiek, čeľuste.
Mesoderm- kostrové svaly, bránica, stavce, dentín, obličkové tubuly, močovody, vajcovody, maternica, časť vaječníkov a semenníkov, kôra nadobličiek, srdce, krv, lymfatický systém, pľúca, skléra, cievnatka a rohovka oka .
Endoderm- notochord, väčšina tráviaceho traktu, črevná výstelka, močového mechúra, pľúca, pankreas, týmus, štítna žľaza, prištítna žľaza.
Pod akordom je črevná trubica, po stranách akordu - somitný mezoderm.
1. Vonkajšia časť somitu, susediaca s ektodermou, je tzv dermotóm. Tvorí spojivové tkanivo pokožky.
2. Interiér - sklerotóm- dáva vznik kostre.
3. Medzi dermotómom a sklerotómom je myotómčím vznikajú priečne pruhované svaly.
4. Pod somitmi sú nohy ( nefragonotóm), z ktorých sa tvorí urogenitálny systém.
Coelomic tašky sú po stranách vytvorené symetricky. Steny coelomových vakov smerujúce do čriev sa nazývajú splanchnopleura, smerom k ektodermu somatopleura. Tieto listy sa podieľajú na vzdelávaní kardiovaskulárny systém, pohrudnica, peritoneum, osrdcovník.
dozorných orgánov.
dočasné orgány- sú to dočasné orgány potrebné pre život embrya. Čas ich vzniku závisí od vajíčka a podmienok prostredia.
Prítomnosť alebo neprítomnosť dočasných orgánov je základom rozdelenia stavovcov do skupín: Amniota a Anamnia.
Do skupiny anamniev zahŕňajú evolučne staršie živočíchy, ktoré sa vyvíjajú vo vodnom prostredí a nepotrebujú ďalšiu vodu a iné schránky embrya.(Cyklostómy, ryby, obojživelníky)
Do skupiny amniote zahŕňajú primárne suchozemské stavovce, ktorých embryonálny vývoj prebieha v suchozemských podmienkach. (plazy, vtáky, cicavce)
V štruktúre a funkciách dočasných orgánov amniotov je veľa spoločného. Provizórne orgány vyšších stavovcov sa nazývajú zárodočné membrány. Vyvíjajú sa z bunkového materiálu už vytvorených zárodočných vrstiev.
Dočasné orgány:
Amnion- vačok naplnený plodovou vodou, ktorý vytvára vodné prostredie a chráni embryá pred vysychaním a poškodením.
Chorion- vonkajšia zárodočná membrána susediaca so schránkou alebo materskými tkanivami. Slúži na výmenu s okolím, podieľa sa na dýchaní, výžive a vylučovaní.
Žĺtkový vak- podieľa sa na výžive embrya a je krvotvorným orgánom.
Alantois- výrastok zadného čreva sa podieľa na výmene plynov, je schránkou pre močovinu a kyselinu močovú. U cicavcov to spolu s chorionom tvorí placentu.Od alantoisu po chorion vyrastajú cievy, ktorými placenta plní vylučovacie, dýchacie a nutričné funkcie.
Druhy placenty:
1.Epiteliochorálna- (semiplacenta) má najjednoduchšiu štruktúru. Keď sa vytvorí, na povrchu chorionu sa objavia klky vo forme malých tuberkulóz, ktoré sa ponoria do zodpovedajúcich vybraní sliznice maternice bez toho, aby ju narušili. (chorion je v kontakte s epitelom žliaz maternice) Konské ošípané
2. Desmochoriový- charakterizovaný vytvorením najužšieho spojenia medzi chorionom embrya a stenou maternice. V mieste kontaktu s vilmi chorionu je epitel zničený. Rozvetvené platničky sú ponorené do spojivového tkaniva. (Chorion je v kontakte so spojivovým tkanivom.)
3. Endotelochorálna- ničí sa nielen epitel, ale aj spojivové tkanivo. Klky sú v kontakte s cievami a od materskej krvi ich oddeľuje len tenká endotelová stena.(predátori)
4. Hemochoriálny- v maternici sú hlboké zmeny. Klky sa kúpajú v krvi a absorbujú z nej živiny.
Autor: vzhľad:
1.Difúzne- Klky sú rovnomerne rozložené po celej ploche chorionu.
2. Cotyledonnaya- klky sa zhromažďujú v skupinách vo forme kríkov
3.Pás- klky tvoria pás obklopujúci vodný mechúr.
4.Diskoidný– Klky sa nachádzajú v diskoidnej oblasti na povrchu chorionu.
Blastula, nazývaná aj zárodočný vak, je konečným výsledkom štiepenia vajíčka. Ďalším stupňom, ktorý zaujíma medzipolohu medzi drvením a organogenézou, v embryogenéze je gastrulácia. Jeho hlavným významom je vytvorenie troch zárodočných vrstiev: endoderm, ektoderm a mezoderm. Inými slovami, gastruláciou sa začína embryonálna diferenciácia a morfogenéza organizmu.
Definícia pojmu "gastrulácia"
Už v roku 1901 bola gastrulácia opísaná ako dráha, ktorou mezodermálne, endodermálne a ektodermálne bunky vstupujú do embrya. Táto definícia predpokladá prítomnosť špeciálnych orgánovotvorných priestorov v blastule. Po pochopení tohto pomerne jednoduchého popisu je ľahké prejsť ku komplexnejšiemu, modernejšiemu významu tohto pojmu. Gastrulácia je sled morfogenetických pohybov, ktorých výsledkom je pohyb rudimentov tkaniva na miesta na ne určené v súlade s „plánom“ organizácie organizmu. Proces je zložitý, zmeny sú sprevádzané rastom a rozmnožovaním, riadeným pohybom a diferenciáciou buniek.
Ak vezmeme do úvahy gastruláciu vo všeobecnejšom zmysle, možno ju definovať ako medzistupeň patriaci k jedinému dynamickému procesu, počas ktorého dochádza k preskupovaniu sekcií blastuly, čo značne uľahčuje prechod k procesu organogenézy.
Pohyb buniek
Ak dáva všeobecné charakteristiky považovaný za proces, potom môžeme povedať, že gastrulácia je embólia a epibólia. Oba pojmy odrážajú morfogenetický pohyb buniek, ku ktorému dochádza v absolútne všetkých štádiách ontogenetického vývoja organizmu. Najvýraznejšie sú však pri gastrulácii. Epibólia je proces pohybu buniek po povrchu embrya a embólia je ich pohyb do neho.
V embryológii sa rozlišujú tieto hlavné typy gastrulácie alebo pohybu buniek: invaginácia, imigrácia, involúcia, delaminácia a epibólia. Viac podrobností o nich - neskôr v článku.
Pohyb bunkových listov
Na procese gastrulácie sa môžu zúčastniť nielen jednotlivé (voľne migrujúce) bunky, ale aj celé bunkové vrstvy. Smer je určený konštantnými a vzdialenými interakciami. Prvé sily objavil P. Weiss v 20. rokoch minulého storočia a zrejme sa vyskytujú v embryogenéze, druhé – zriedkavé a špeciálne, sa vyskytujú s malou mierou pravdepodobnosti pri normálnej morfogenéze.
Počas gastrulácie nedochádza k fragmentácii buniek. Ako bolo uvedené vyššie, začína sa pohyb bunkových hmôt a v dôsledku toho sa vytvára dvojvrstvové embryo, nazývané gastrula. Endoderm a ektoderm sú jasne viditeľné. Vo všetkých mnohobunkových organizmoch (výnimkou sú len coelenteráty) sa súbežne s gastruláciou alebo bezprostredne po nej vytvára tretia zárodočná vrstva, nazývaná mezoderm. Je to súbor buniek umiestnených medzi ektodermou a endodermou. V dôsledku toho sa embryo stáva trojvrstvovým.
Metódy gastrulácie priamo závisia od typu blastuly.
Invaginovaná gastrula

Názov metódy hovorí sám za seba. Invaginácia je invaginácia jednovrstvovej steny blastuly (balstodermu) do blastocoelu. Najprimitívnejší a najnázornejší príklad bude s gumenou loptičkou. Keď ho stlačíte, časť materiálu sa stlačí dovnútra. Invaginácia môže byť privedená k najvzdialenejšej stene alebo bezvýznamná. V dôsledku toho sa blastula transformuje a gastrula sa získa vo forme dvojvrstvového vrecka s archenteronom. Jeho vnútorná stena je primárny endoderm a jeho vonkajšia stena je primárny ektoderm. Vzniknutý archenterón (primárne črevo) komunikuje s vonkajším prostredím cez otvor nazývaný blastopór. Jeho druhé meno je primárne ústa. Jeho ďalší vývoj závisí od typu organizmu. U mnohých zvierat sa z blastopóru nakoniec vyvinú definitívne ústa. V tomto ohľade sa nazývajú protostómy (mäkkýše, červy, článkonožce). V deuterostómoch sa blastopór mení na neurointestinálny kanál umiestnený v zadnej časti embrya (v strunatách) alebo v konečníku.
imigračná gastrula

Imigračná gastrulácia je metóda tvorby dvojvrstvového embrya, najcharakteristickejšieho z coelenterátov. Gatrula vzniká aktívnym vysunutím časti buniek blastuly do blastocoelu. Takáto imigrácia je unipolárna. Bunky sa pohybujú iba z vegetatívneho pólu. Neskôr tvoria endoderm, teda vnútornú vrstvu. Týmto spôsobom sa gastrulácia uskutočňuje v hydroidnom polype, medúze.
Blastodermálne bunky môžu preniknúť do blastocoelu nie v jednej oblasti, ale po celom povrchu embrya. Takáto imigrácia sa nazýva multipolárna, ale je pomerne zriedkavá.
V mnohých coelenterátoch, ktoré sa vyznačujú imigračným spôsobom gastrulácie, dochádza k veľmi aktívnemu „vysťahovaniu“ buniek blastuly a vzniknutá gastrula úplne stráca blastocoel. V tomto prípade chýba blastopór charakteristický pre predchádzajúcu metódu invaginácie.
Delaminačná gastrula

The vzácny pohľad gastrula bola prvýkrát opísaná Mečnikovom I.I. a je charakteristická pre coelenteráty. Procesy sprevádzajúce gastruláciu sú veľmi zvláštne, ale pri zvažovaní typického prípadu sú vnímané jednoduchšie. Napríklad vajíčka niektorých scyphomedusa majú koncentricky umiestnené a dobre rozlíšiteľné časti cytoplazmy: husté a granulárne (ektoplazma) a bunkové (endoplazma). Vyznačujú sa relatívne synchrónnym a rovnomerným delením: 2, 4, 8, 16. V konečnom dôsledku embryo obsahuje 32 blastomérov. Ďalšie delenie sa vykonáva rovnobežne s povrchom embrya. Vytvára sa vonkajšia vrstva blastomérov, pozostávajúca z ektoplazmy a vnútorná vrstva, čiastočne z ektoplazmy a endoplazmy. Inými slovami, proces tvorby viacvrstvového embrya prebieha rozdelením jednej vrstvy buniek na dve. Potom sú rozdrvené iba vnútorné blastoméry a opäť rovnobežné s povrchom embrya, ktoré v dôsledku takejto zvláštnej gastrulácie nadobúda tvar gule. Skladá sa zo 64 plochých buniek, ktoré tvoria ektodermu a 32 konvexnejších buniek, ktoré sú základom endodermy.
epibolická gastrula

U zvierat s výraznou telolecitálnou štruktúrou vajíčok (presun žĺtka na vegetatívny pól) dochádza k gastrulácii podľa epibolickej metódy. Makroméry sú veľké blastoméry, ktoré sa delia veľmi pomaly a obsahujú veľké množstvo žĺtka. Nemajú schopnosť pohybu, v súvislosti s tým sa po nich aktívnejšie mikroméry nachádzajúce sa na povrchu bunky doslova „plazia“. Pri takejto gastrulácii chýba blastopór a netvorí sa archenterón. Až v budúcnosti, keď sa makroméry napriek tomu zmenšia, začne sa vytvárať dutina, základ primárneho čreva.
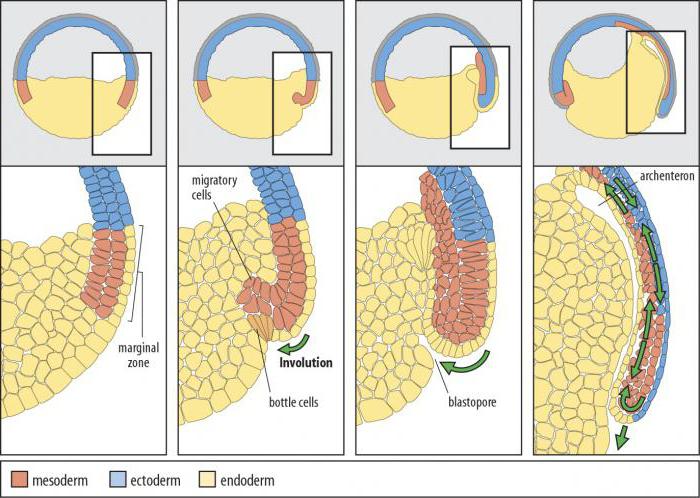
involúcia
Involučná gastrulácia je proces, ktorý spočíva v „zastrčení“ vonkajšej vrstvy buniek do vnútra embrya. Zväčšuje sa a šíri sa pozdĺž vnútorného povrchu. Tento spôsob gastrulácie je charakteristický pre zvieratá s mezolecitálnymi vajíčkami - obojživelníky (obojživelníky). Pohyb vedúcich hlbokých buniek okrajovej oblasti inhibuje vývoj archenterónu. Práve v nich je hybná sila involúcie.
Zmiešaný spôsob gastrulácie
Ako viete, embryogenéza je najviac skoré obdobie vývoj každého jednotlivého organizmu: od počatia po narodenie. Gastrulácia je jedným z jeho štádií, druhým v chronológii po rozdrvení. Jej metódy sú natoľko odlišné, že ich možno porovnávať s vysokou mierou konvenčnosti. Každý z nich si vyžaduje podrobné štúdium a analýzu. Stále však medzi nimi existujú určité priesečníky. Takže ako druh variantu invaginácie možno považovať proces epiboly a delaminácia má podobnosť s imigráciou.
Všimnite si, že u mnohých zvierat dochádza k gastrulácii kombinovaným spôsobom. V takýchto prípadoch epibólia a invaginácia, ako aj iné morfogenetické procesy prebiehajú súčasne. Najmä takto sa gastrulácia vyskytuje u obojživelníkov. V tomto ohľade mnohí autori rozlišujú zmiešanú metódu.
gastrula

Doslova z latinčiny sa výraz „gastrula“ prekladá ako „lono, žalúdok“. Označuje špecifický zárodok mnohobunkových organizmov. Charakteristickým znakom gastruly je prítomnosť dvoch alebo troch zárodočných vrstiev. Proces jeho tvorby je fázou gastrulácie.
Najjednoduchšie zariadenie sa pozoruje u zvierat. Vyznačujú sa elipsoidnou gastrulou s jednobunkovou vonkajšou vrstvou (ektodermou) a vnútornou akumuláciou buniek (endodermou), ako aj „primárnym črevom“. Za typickú sa považuje gastrula ježovky, ktorá vzniká invagináciou. U ľudí prebieha gastrulácia na 8. – 9. deň vývoja. Gatrula je diskovitý sploštený útvar vytvorený z vnútornej bunkovej hmoty.
U väčšiny zvierat v štádiu gastruly embryo spravidla nemôže žiť voľne a nachádza sa v maternici alebo vo vaječných membránach. Existujú však aj výnimky. Takže larvy črevných dutín, planula, sú voľne plávajúce gastruly.
γαστήρ - žalúdok, maternica) - štádium embryonálneho vývoja mnohobunkových zvierat, ktoré nasleduje po blastule. Výrazná vlastnosť gastrula je tvorba takzvaných zárodočných vrstiev – vrstiev (vrstiev) buniek. V črevných dutinách sa v štádiu gastruly tvoria dve zárodočné vrstvy: vonkajšia je ektoderm a vnútorná je endoderm. V iných skupinách mnohobunkových živočíchov sa v štádiu gastruly vytvárajú tri zárodočné vrstvy: vonkajší - ektoderm, vnútorný - endoderm a stredný - mezoderm. Proces vývoja gastruly sa nazýva gastrulácia.Najjednoduchšie usporiadaná gastrula sa nachádza v Coelenterate - je to embryo elipsoidného tvaru, v ktorom je ektoderm reprezentovaná vonkajšou jednobunkovou vrstvou a endoderm je vnútorná akumulácia buniek. Vo vnútornej vrstve embrya (endoderme) sa vytvorí dutina – tzv. „primárne črevo“ alebo gastrocoel. Neskôr, na prednom konci embrya, tzv. „Primárne ústa“ alebo blastopór je otvor, cez ktorý primárne črevo komunikuje s vonkajším prostredím.
Za typickú gastrulu sa považuje gastrula ježoviek. Vzniká „invagináciou“ časti povrchu sférickej blastuly. V dôsledku intususcepcie je časť blastodermu (koža blastuly) zatlačená dovnútra a tvorí gastrocoel (primárne črevo). Gastrocoelové bunky patria do endodermy. Časť blastodermu zostáva na povrchu embrya a tvorí ektodermu. Časť buniek „je vysťahovaná“ v priestore medzi vonkajšou vrstvou embrya a primárnym črevom, tieto bunky tvoria mezoderm. Taktiež z primárneho čreva vo vnútri embrya sú oddelené tzv. coelomové vaky, ktoré sú tiež súčasťou mezodermu. Otvor, cez ktorý dochádza k invaginácii, sú primárne ústa (blastopór).
Ľudské embryo prechádza štádiom gastruly na 8. – 9. deň vývoja. Ľudská gastrula je sploštený diskoidný útvar (takzvaný „zárodočný disk“), ktorý sa tvorí z „vnútornej bunkovej hmoty“ blastocysty. Horná (to znamená privrátená k zvieraciemu pólu) vrstva zárodočného disku sa označuje ako ekdoterm, stredná vrstva sa označuje ako mezoderm, spodná (to znamená otočená k vegetatívnemu pólu k budúcemu žĺtkovému vaku). ) vrstva disku sa označuje ako endoderm. Homológom primárneho čreva u človeka je tzv. „primárny žĺtkový vak“ – priestor ohraničený od zvieracieho pólu ektodermou zárodočného disku, a z ostatných strán tzv. hypoblastóm - extraembryonálny endoderm.
Gatrula sa môže vytvoriť invagináciou (invagináciou alebo embolizáciou) alebo epibóliou (napríklad u niektorých bezstavovcov). Pri epibolii malé ektodermálne bunky postupne prerastajú veľké endodermálne bunky, pričom dutina sa nevytvorí okamžite, ale objaví sa neskôr.
U väčšiny zvierat embryo v štádiu gastruly nie je voľne žijúce, ale nachádza sa vo vaječných membránach alebo v maternici. Existujú však zvieratá s voľne plávajúcimi gastrulami (napríklad voľne plávajúca larva Coelenterates - planula (parenchymula) - je gastrula).
Evolučný pôvod gastruly
Prítomnosť štádia gastruly u všetkých mnohobunkových živočíchov je jedným z dôkazov jednoty ich pôvodu. Podľa Haeckelovho-Mullerovho biogenetického zákona táto okolnosť naznačuje spoločného predka, ktorý existoval u všetkých mnohobunkových živočíchov a svojou štruktúrou pripomínal gastrulu moderných živočíchov. Existuje niekoľko hypotéz týkajúcich sa evolučného pôvodu tohto hypotetického predchodcu metazoanov podobného gastrule.
Ernst Haeckel v roku 1872 predložil tzv. "Teória Gastrea". Podľa tejto hypotézy boli predkami všetkých mnohobunkových organizmov guľovité mnohobunkové kolónie bičíkovcov (podobne ako blastula Haeckel nazval tento pôvodný organizmus „blastea“), ktoré plávali v mori ako súčasť planktónu a živili sa malými organickými časticami suspendovanými vo vode (napríklad baktérie). V priebehu evolúcie blastea podstúpila invagináciu (invagináciu) a vytvorila organizmus pozostávajúci z dvoch vrstiev buniek (vonkajšia a vnútorná), vnútorná vrstva buniek tvorila „črevo“, ktoré sa otváralo von „ústami“ (napr. podobnosť s gastrulou, Haeckel nazval tento rodový organizmus „gastreya“). Podľa E. Haeckela biologický význam premeny blastea na gastreu spočíval v špecializácii buniek. Všetky bunky blastea boli rovnaké; pomocou bitia bičíkov bunky podporovali blastea vo vodnom stĺpci a tiež naberali častice potravy na prehltnutie. Špecializácia nastala na gastreu: bunky vonkajšej vrstvy pomocou úderu bičíka podopierali gastreu vo vodnom stĺpci, bunky vnútornej vrstvy pomocou úderu bičíka vytvorili tzv. prietok tekutiny, ktorý vtiahol častice do primárneho čreva. Prítomnosť dutiny v gastrea jej poskytla evolučnú výhodu - gastrea, na rozdiel od blastea, mala schopnosť jesť potravu, ktorej veľkosť je väčšia ako bunky samotnej gastrey, pretože teraz bunky vnútornej vrstvy môžu vylučovať tráviace enzýmy do žalúdočnej dutiny. Podľa teórie gastrulácie je najstarším typom gastrulácie invaginácia, ostatné typy gastrulácie sú sekundárne a objavili sa neskôr vo vývoji. Najprimitívnejšia forma gastruly, planula, je teda sekundárne zjednodušená embryonálna forma živočíchov.
Iľja Iľjič Mečnikov v rokoch 1876-1886 sformuloval tzv. teória fagocytely. Podľa tejto hypotézy evolúcia blastea neprebiehala invagináciou, ale vysunutím buniek vonkajšej vrstvy vo vnútri sférickej blasty. Mečnikov zdôvodnil takéto vysťahovanie ("imigráciu") nasledovne: bunky blastea po zachytení častíc potravy (fagocytóza) boli oddelené od vonkajšej vrstvy a ponorené do blastea na strávenie. Na konci trávenia boli bunky reintegrované do vonkajšej vrstvy. Tento proces prebiehal nepretržite. Mechnikov nazval tento hypotetický staroveký organizmus "phagocytella" alebo "paranchimella". V prospech fagocytárnej teórie hovorí skutočnosť, že najprimitívnejšie mnohobunkové živočíchy vytvárajú gastrulu imigráciou buniek vonkajšej vrstvy dovnútra, ako aj skutočnosť, že najjednoduchšie usporiadané mnohobunkové živočíchy nemajú trávenie dutín, ale iba vnútrobunkové. . Podľa teórie fagocytely je najstarším typom gastrulácie imigrácia. Slabým miestom fagocytárnej teórie je, že nevysvetľuje biologický význam migrácie fagocytárnych buniek do kolónie.
pozri tiež
Literatúra
Nadácia Wikimedia. 2010.
Synonymá:Pozrite sa, čo je „Gastrula“ v iných slovníkoch:
Jedna z embryonálnych foriem embrya zvierat. Slovník cudzích slov zahrnutých v ruskom jazyku. Chudinov, A.N., 1910. GASTRULA je špeciálna forma, štádium vývoja, ktorým prechádza embryo zvieraťa. Kompletný slovník cudzích slov zahrnutý v ... ... Slovník cudzích slov ruského jazyka
- (novolat. gastrula) štádium embryonálneho vývoja mnohobunkových živočíchov nasledujúce po blastule. Gatrula má dve, potom trojvrstvovú stenu a dutinu (gastrocoel), zvyčajne komunikujúcu s vonkajším prostredím blastopórom ... Veľký encyklopedický slovník
GASTRULA, rané štádium vývoja zvieracích embryí. Predchádza mu BLASTULA stage. Gatrula je dutina s dvoma vrstvami (pozri VRSTVA GEM) buniek. Vnútorná vrstva je ENDODERM, vonkajšia vrstva je ECTODERM. Dutina embrya sa nazýva gastrocoel a jeho ... ... Vedecko-technický encyklopedický slovník
Konvexnosť, zárodok Slovník ruských synoným. gastrula n., počet synoným: 2 konvexita (41) ... Slovník synonym
- (z gréckeho gaster žalúdok), zárodok mnohobunkových živočíchov pri gastrulácii. G. bol prvýkrát opísaný A. O. Kovalevským v roku 1865 a pomenovaný ako "črevná larva", termín "G." zaviedol v roku 1874 E. Haeckel. Zvyčajne existujú štádiá skorého, stredného a neskorého G ... Biologický encyklopedický slovník
GASTRULA- (z gréckeho gaster žalúdok), embryologický termín zavedený Haeckelom na označenie 3. štádia vývoja nasledujúceho po štádiu blastuly (pozri); proces tvorby G. sa nazýva gastrulácia. Embryo v G. štádiu má dve ... ... Veľká lekárska encyklopédia
gastrula- Embryo mnohobunkového živočícha počas gastrulácie, ktoré má tri hlavné zárodočné vrstvy – ektoderm, endoderm (okrem húb a čriev) a mezoderm; G. prvýkrát opísal A.O. Kovalevského v roku 1865 a termín ... ... Technická príručka prekladateľa
- (novolat. gastrula), štádium embryonálneho vývoja mnohobunkových živočíchov nasledujúce po blastule. Gatrula má dve, potom trojvrstvovú stenu a dutinu (gastrocoel), zvyčajne komunikujúcu s vonkajším prostredím blastopórom. * * * GASTRULA GASTRULA… … encyklopedický slovník
Gastrula gastrula. Embryo mnohobunkového zvieraťa počas gastrulácie
GASTRULA- (gastrula) skoré štádium embryonálneho vývoja u mnohých zvierat. Gatrula je dvojvrstvová stena a centrálna dutina archenterónu, ktorá sa otvára smerom von cez blastopór. Skutočná gastrulácia sa vyskytuje iba ... ... Výkladový slovník medicíny
Za procesy štiepenia a blastulácie nasleduje celý rad ďalších vývojových procesov, ktoré vedú k vytvoreniu primitívnych, primárnych anláží orgánov embrya, teda k vzniku najskôr dvoch a potom troch zárodočných vrstiev alebo vrstiev z pôvodne jedinej blastodermy zárodku. blastula (vonkajšia zárodočná vrstva alebo ektoderm, stredný list alebo mezoderm a vnútorný list alebo endoderm). Štádium vývoja s dvoma základnými zárodočnými vrstvami sa nazýva gastrula.
Po skončení obdobia gastrulácia dochádza už k zložitejším zmenám, ktoré najskôr spôsobujú vznik chrbtovej časti embrya (notogenéza), neskôr - vznik jeho tela, uloženie telovej dutiny, tzv. coelom v mezoderme (coelomatizácia), potom kladenie chrbtovej struny, teda struny (chordulácia), a zárodkov tvorby nervový systém vo forme neurálnej, medulárnej platničky a medulárnej trubice (neurulácia). Čím je živočích fylogeneticky starší, teda čím vyššie je štádium jeho fylogenetického vývoja, tým sú tieto procesy zložitejšie (hoci ich základná schéma zostáva nezmenená a možno ju odvodiť z vývoja lanceletu) a tým viac sa tieto procesy chronologicky prekrývajú ( heterochronicky).
V oblasti vegetačného pólu blastula v blastoderme o niečo väčšie ako blastoméry na zvieracom póle. Oblasť týchto väčších blastomér vegetatívneho pólu sa začína postupne vtláčať, invaginovať do blastocoelu smerom k zvieraciemu pólu.
Z tohto dôvodu dutina blastocoel začína klesať a blastoderm vegetatívneho pólu sa približuje k bunkovej vrstve blastodermy zvieracieho pólu. Nakoniec obe tieto vrstvy na seba nadväzujú, čo súčasne vedie k zániku blastocoelovej dutiny. Tento proces možno čisto morfologicky porovnať s vtlačením jednej steny perforovanej gumovej gule dovnútra, smerom k protiľahlej stene. Môžeme povedať, že gastrulácia v lancelete prebieha podľa metódy intususcepcie (invaginácie).
Neexistuje žiadny aktívny zarastený oblasti vegetatívneho pólu smerom k zvieraciemu pólu; proces invaginácie je v skutočnosti spôsobený nerovnomerným rastom blastuly. Tým, že bunky v oblasti zvieracieho pólu sa množia rýchlejšie ako bunky na vegetatívnom póle, blastoderm zvieracieho pólu, ktorý rastie do šírky, sa začína uzatvárať, vrátane pomalšie rastúcej oblasti vegetatívnych blastomér.
Spolu s, bezpochýb, dôležité sú aj zmeny v koloidnom stave povrchovej vrstvy cytoplazmy buniek pozdĺž okrajov blastopóru.
Teda ako výsledok invaginácie objavuje sa miskovitý útvar, ktorého stena je už dvojvrstvová, keďže protiľahlé oblasti blastodermu pri invaginácii k sebe priliehajú. Na dne novej dutiny, ktorá je výsledkom invaginácie, je vnútorná vrstva buniek zodpovedajúca bývalému vegetatívnemu pólu blastuly. Jeho vonkajší povrch je naopak pokrytý vrstvou blastodermu, ktorá sa predtým nachádzala na zvieracom póle. Toto štádium vývoja, ktoré je charakterizované stenou pozostávajúcou z dvoch susediacich bunkových vrstiev, sa nazýva gastrula. Vonkajšia epiteliálna vrstva gastruly je vonkajšia zárodočná vrstva, vrstva je ektoderm, vnútorná vrstva je vnútorná zárodočná vrstva - endoderm.
Podstata štádia gastrulácie spočíva v tom, že jednovrstvové embryo – blastula – sa mení na viacvrstvové - dvoj- alebo trojvrstvové, tzv gastrula(z gréčtiny. gaster -žalúdok v zdrobnenom význame).
U primitívnych strunatcov, napríklad v lancelete, sa homogénna jednovrstvová blastoderm počas gastrulácie premení na vonkajšiu zárodočnú vrstvu - ektodermu - a vnútornú zárodočnú vrstvu - endoderm. Endoderm tvorí primárne črevo s dutinou vo vnútri gastrocoel. Otvor vedúci ku gastrocoelu je tzv blastopor alebo primárne ústa. Dve zárodočné vrstvy definujú morfologické znaky gastrulácie. Ich existencia v určitom štádiu vývoja u všetkých mnohobunkových živočíchov, od koelenterátov až po vyššie stavovce, nám umožňuje uvažovať o homológii zárodočných vrstiev a jednote pôvodu všetkých týchto živočíchov.
U stavovcov sa pri gastrulácii okrem spomínaných dvoch vytvára ešte tretia zárodočná vrstva - mezoderm, nachádza sa medzi ekto- a endodermou. Vývoj strednej zárodočnej vrstvy, ktorou je chordomesoderm, je evolučnou komplikáciou gastrulačnej fázy u stavovcov a je spojený s urýchlením ich vývoja skoré štádia embryogenéza. U primitívnejších strunatcov, ako je lancelet, sa chordomesoderm zvyčajne vytvára na začiatku fázy nasledujúcej po gastrulácii - organogenéza. Posun v čase vývoja niektorých orgánov v porovnaní s inými u potomkov v porovnaní so skupinami predkov je prejavom heterochrónia. Zmeny v načasovaní formovania najdôležitejších orgánov v priebehu evolúcie nie sú nezvyčajné.
Charakteristický je proces gastrulácie dôležité bunkové premeny, ako sú riadené pohyby skupín a jednotlivých buniek, selektívne rozmnožovanie a triedenie buniek, začiatok cytodiferenciácie a indukčné interakcie. Uvedené bunkové mechanizmy ontogenézy sú podrobne diskutované v kap. 8.2.
Ryža. 7.3. Predpokladané základy, gastrulácia a neurulácia v lancelete.
ALE - predpokladané rudimenty v štádiu blastuly (vonkajší pohľad) a ranej gastruly (rezový pohľad); B - neskorá gastrula a neurulácia v sagitálnych (ľavý rad) a priečnych (pravý rad) rezoch; AT - plastický model embrya na konci neurulačného obdobia:
1- zvieracia palica, 2- vegetatívny pól, 3- blastocoel, 4- gastrocoel, 5-dorzálne a ventrálne pery blastopóru, 6 - hlavový koniec embrya, 7-modulárna doska, 8 - chvostový koniec embrya, 9-dorzálna časť mezodermu, 10- dutina sekundárneho čreva. 11 - segmentované somity, 12- brušná časť mezodermu; a, b, c, d, e - označenie predpokladaných a vyvíjajúcich sa orgánov: a- ektodermová koža, b - nervová trubica, v - akord, G - endoterma, črevný epitel, d - mezodermom
Gastrulačné metódy rôzne. Rozlišujú sa štyri typy priestorovo usmernených pohybov buniek, ktoré vedú k premene embrya z jednej vrstvy na viacvrstvovú.
invaginácia - invaginácia jedného z úsekov blastodermu dovnútra ako celá vrstva. V lancelete invaginujú bunky vegetatívneho pólu, u obojživelníkov dochádza k intususcepcii na hranici medzi zvieracím a vegetatívnym pólom v oblasti sivého polmesiaca. Proces invaginácie je možný iba vo vajciach s malým alebo stredným množstvom žĺtka.
Epiboly - zanášanie malými bunkami živočíšneho pólu väčších, zaostávajúcich v rýchlosti delenia a menej pohyblivých buniek vegetatívneho pólu. Tento proces je jasne vyjadrený u obojživelníkov.
Denominácia - stratifikácia blastodermových buniek do dvoch vrstiev ležiacich nad sebou. Delamináciu možno pozorovať v diskoblastule embryí s čiastočným typom drvenia, ako sú plazy, vtáky a vajcorodé cicavce. Delaminácia sa prejavuje v embryoblastoch placentárnych cicavcov, čo vedie k vytvoreniu hypoblastu a epiblastu.
imigrácia - pohyb skupín alebo jednotlivých buniek, ktoré nie sú spojené do jednej vrstvy. Imigrácia sa vyskytuje u všetkých embryí, ale je najcharakteristickejšia pre druhú fázu gastrulácie u vyšších stavovcov.
V každom konkrétnom prípade embryogenézy sa spravidla kombinuje niekoľko metód gastrulácie.
Morfológia gastrulácie. Podrobnejšie zváženie gastrulácie u lanceletu, žaby, sliepky a cicavcov, na ktoré sa obrátime, pomôže lepšie pochopiť evolučné vzťahy a pochopiť zákonitosti individuálneho vývoja.
gastrulácia lancelet znázornené na obr. 7.3. Rôzne markery v štádiu blastuly (obr. 7.3, A) označené predpokladaný(predpokladané) začiatky. Ide o oblasti blastuly, z ktorých bunkového materiálu sa pri gastrulácii a včasnej organogenéze (neurulácii) zvyčajne vytvárajú úplne definované zárodočné vrstvy a orgány (obr. 7.3, Obr. B a AT).
Invaginácia začína na vegetatívnom póle. Vďaka rýchlejšiemu deleniu bunky živočíšneho pólu rastú a vytláčajú bunky vegetatívneho pólu do blastuly. To je uľahčené zmenou stavu cytoplazmy v bunkách, ktoré tvoria pery blastopóru a priľahlé k nim. V dôsledku invaginácie klesá blastocoel a zvyšuje sa gastrocoel. Súčasne s vymiznutím blastocoelu sa ektoderm a endoderm dostanú do úzkeho kontaktu. V lancelete, ako vo všetkých deuterostómoch (zahŕňajú typ ostnatokožca, typ strunatca a niektoré ďalšie malé druhy živočíchov), sa oblasť blastopóru mení na chvostovú časť organizmu, na rozdiel od protostómov, v ktorých blastopor zodpovedá do hlavovej časti. Ústny otvor v deuterostómoch sa tvorí na konci embrya oproti blastopóru.
Ryža. 7.4. Bunky v tvare banky v oblasti blastopóru skorej gastruly obojživelníkov: 1 - lepidlá v tvare kužeľa, 2 - chrbtová pera
Gastrulácia u obojživelníkov má veľa spoločného s gastruláciou lanceletu, ale keďže žĺtok v ich vajciach je oveľa väčší a nachádza sa hlavne na vegetatívnom póle, veľké blastoméry amfiblastuly sa nedokážu vydúvať dovnútra. Intususcepcia ide trochu inak. Na hranici medzi zvieracím a vegetatívnym pólom v oblasti sivého kosáka sa bunky najskôr silne natiahnu dovnútra, pričom majú tvar „baňky“ (obr. 7.4), a potom vytiahnu bunky povrchovej vrstvy blastula spolu s nimi. Objaví sa polmesiačiková ryha a dorzálny blastopórový pysk.
Súčasne sa menšie bunky zvieracieho pólu, ktoré sa delia rýchlejšie, začnú pohybovať smerom k vegetatívnemu pólu. V oblasti chrbtovej pery sa vytáčajú a invaginujú a väčšie bunky rastú po stranách a na opačnej strane kosákovitej ryhy. Potom proces epiboly vedie k vytvoreniu laterálnych a ventrálnych pyskov blastopóru. Blatopór sa uzatvára do prstenca, vo vnútri ktorého sú nejaký čas viditeľné veľké svetelné bunky vegetatívneho pólu v podobe takzvanej žĺtkovej zátky. Neskôr sú úplne ponorené dovnútra a blastopór sa zužuje.
Pomocou metódy značenia vitálnymi (vitálnymi) farbivami u obojživelníkov boli podrobne študované pohyby buniek blastuly počas gastrulácie.Zistilo sa, že špecifické oblasti blastodermy, tzv. predpokladaný(z lat. praesumptio - predpoklad), pri normálnom vývoji sa objavujú najskôr v zložení určitých rudimentov orgánov, a potom v zložení orgánov samotných (obr. 7.5). Je známe, že u bezchvostých obojživelníkov materiál predpokladaného notochordu a mezodermu v štádiu blastuly neleží na jeho povrchu, ale vo vnútorných vrstvách steny amfiblastuly, avšak približne v úrovniach znázornených na obrázku. Analýza raných štádií vývoja obojživelníkov nám umožňuje dospieť k záveru ovoplazmatická segregácia, ktorý sa zreteľne prejavuje vo vajíčku a zygote (obr. 7.6), má veľký význam pri určovaní osudu buniek, ktoré zdedili ten či onen úsek cytoplazmy. Určitá podobnosť medzi procesmi gastrulácie a oblasťou predpokladaných orgánov u obojživelníkov a lanceletu, t.j. homológia hlavných orgánov, ako je nervová trubica, notochorda, sekundárne črevo, naznačuje ich fylogenetický vzťah.
Ryža. 7.5. Mapa oblastí predpokladaných orgánových primordií v skorých štádiách embryonálneho vývoja obojživelníkov. ALE -štádium blastuly (ochabnuté vľavo); B-D - postupné štádiá gastrulácie (sagitálne rezy); E - začiatok neurulácie (prierez):
1 - kožný ektoderm 2- nervová trubica, 3- notochord, 4-mezoderm somitov, 5- mezoderm splanchnotómov, 6 - endoderm, 7 - blastocoel, 8 - polmesiaca drážka, 9-gastrocoel, 10- blastopór dorzálnej pery, 11 - žĺtka 12- dutina sekundárneho čreva 13- nervové záhyby
Gastrulácia v embryách s meroblastickým typom štiepenia a vývoja má svoje vlastné charakteristiky. O vtákov začína po rozštiepení a vytvorení blastuly pri prechode embrya cez vajcovod. V čase, keď je vajíčko znesené, embryo už pozostáva z niekoľkých vrstiev: horná vrstva sa nazýva epiblast, nižšie - primárny hypoblastóm(obr. 7.2, AT). Medzi nimi je úzka medzera - blastocoel. Potom sa vytvoril sekundárny hypoblast, spôsob vzniku nie je celkom jasný. Existujú dôkazy, že primárne zárodočné bunky pochádzajú z primárneho hypoblastu vtákov, zatiaľ čo sekundárne tvoria extraembryonálny endoderm. Vznik primárneho a sekundárneho hypoblastu sa považuje za jav predchádzajúci gastrulácii.
Hlavné udalosti gastrulácie a konečnej tvorby troch zárodočných vrstiev sa začínajú po kladení vajíčok so začiatkom inkubácie. Dochádza k hromadeniu buniek v zadnej časti epiblastu v dôsledku nerovnomernej rýchlosti bunkového delenia a ich pohybu z laterálnych častí epiblastu do stredu, smerom k sebe. Takzvaný primárna línia, ktorý sa tiahne smerom k hlavovému koncu. V strede primárneho pruhu sa tvorí primárna brázda, a pozdĺž okrajov - primárne valčeky. Na hlavovom konci primárneho prúžku sa objaví zhrubnutie - Hensenov uzol, a v ňom - primárna fossa (obr. 7.7).
Keď bunky epiblastu vstúpia do primárnej drážky, ich tvar sa zmení. Tvarom sa podobajú na bunky „baňovitého tvaru“ gastruly obojživelníkov. Tieto bunky sa potom stanú hviezdicovitými a ponoria sa pod epiblast za vzniku mezodermu (obr. 7.8). Endoderm sa tvorí na báze primárneho a sekundárneho hypoblastu s pridaním novej generácie endodermálnych buniek migrujúcich z horných vrstiev, blastodermy. Prítomnosť niekoľkých generácií endodermálnych buniek naznačuje predĺženie doby gastrulácie v čase.
Ryža. 7.6. Ovoplazmatická segregácia vo vajciach obyčajných žiab.
ALE - ihneď po oplodnení; B- 2 hodiny po oplodnení (pohľad zľava): 1 - pigmentovaná oblasť zvieraťa, 2- nepigmentovaná negatívna oblasť, 3 - os hlava-chvost budúceho organizmu, 4- sivý kosák, 5 - chrbtová strana, 6 - ventrálna strana
Ryža. 7.7. Kuracie embryo v štádiu primárneho pruhu
(pohľad zozadu):
1 - tmavá oblasť, 2 - priesvitná oblasť zárodočného disku
Časť buniek migrujúcich z epiblastu cez Hensenov uzol tvorí budúci notochord. Súčasne s iniciáciou a predĺžením chordy postupne mizne Hensenov uzol a primárny pruh v smere od predného ku kaudálnemu koncu. Tomu zodpovedá zúženie a uzavretie blastopóru. Keď sa primárny pruh zmršťuje, zanecháva za sebou vytvorené časti osových orgánov embrya v smere od hlavy k častiam chvosta. Zdá sa rozumné považovať pohyby buniek v kuracom embryu za homológne epiboly a primárny pruh a Hensenov uzol za homológne s blastopórom v dorzálnej pere gastruly obojživelníka.
Je zaujímavé poznamenať, že bunky embryí cicavcov (kapitola 7.6.1), napriek skutočnosti, že u týchto zvierat majú vajíčka malé množstvo žĺtka a fragmentácia je úplná, vo fáze gastrulácie si zachovávajú pohyby charakteristické pre embryá plazov a vtákov. To potvrdzuje myšlienku pôvodu cicavcov zo skupiny predkov, ktorých vajcia boli bohaté na žĺtok.
Ryža. 7.8. Kuracie embryo v štádiu primárneho pruhu (prierez).
A, B - pri malom a veľkom zväčšení: 1 - ektoderm, 2 - endoderm, 3 - mezoderm, 4 - primárny valec, 5 - primárna drážka
Vlastnosti štádia gastrulácie. Gastrulácia je charakterizovaná rôznymi bunkovými procesmi. Mitotic pokračuje reprodukcia buniek, navyše má v rôznych častiach embrya rôznu intenzitu. Avšak najviac vlastnosť gastrulácia pozostáva z pohyb bunkových hmôt. To vedie k zmene štruktúry embrya a jeho premene z blastuly na gastrulu. deje triedenie bunky svojou príslušnosťou k rôznym zárodočným vrstvám, vo vnútri ktorých sa navzájom „spoznávajú“.
Začína sa fáza gastrulácie cytodiferenciácia,čo znamená prechod k aktívnemu využívaniu biologickej informácie vlastného genómu. Jeden z regulátorov genetickej aktivity je rôzny chemické zloženie cytoplazma embryonálnych buniek, vytvorená ako výsledok ovoplazmatickej segregácie. Takže ektodermálne bunky obojživelníkov majú tmavá farba kvôli pigmentu, ktorý sa do nich dostal zo zvieracieho pólu vajíčka, a bunky endodermu sú svetlé, keďže pochádzajú z vegetatívneho pólu vajíčka.
Počas gastrulácie je úloha veľmi veľká embryonálna indukcia. Ukázalo sa, že výskyt primárneho pruhu u vtákov je výsledkom indukčnej interakcie medzi hypoblastom a epiblastom. Hypoblast má polaritu. Zmena polohy hypoblastu voči epiblastu spôsobuje zmenu orientácie primitívneho pruhu.
Všetky tieto procesy sú podrobne popísané v kapitole 8.2. Treba poznamenať, že tieto prejavy bezúhonnosť ako klíčky determinácia, embryonálna regulácia a integrácia sú jej vlastné počas gastrulácie v rovnakej miere ako počas drvenia (pozri časť 8.3).







